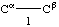
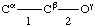
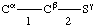
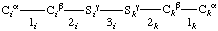
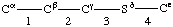
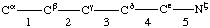
Continued from 3. The Main Chain (or Polypeptide Backbone)
Contents
Atoms are lettered, or lettered and numbered, from Cα, and bonds are numbered from Cα, working outwards away from the main chain.
4.2. Designation of atoms other than hydrogen.
Atoms other than hydrogen are designated in the usual way by Greek letters, β, γ, δ etc., e.g., Ciβ (or Cβ(i )), Niζ (or Nζ(i )).
Note.
The notations for tbe amino acids normally occurring in proteins are given in Table III.
Table III: Symbols for Atoms and Bonds in the Side Chains of the Commonly Occurring L-Amino Acids.
| (a) Unbranched side chains | ||||
|---|---|---|---|---|
| Alanine | | | Serine | |
| Cysteine | | Cystine | | |
| Methionine | | Lysine | | |
| (b) Branched side chains | |||||
|---|---|---|---|---|---|
| Valine | 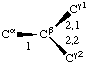 | | Threonine | 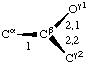 | |
| Isoleucine | 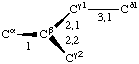 | Leucine | 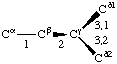 | ||
| Aspartic acid | 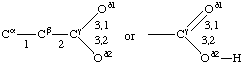 | Asparagine | 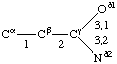 | ||
| Glutamic acid | 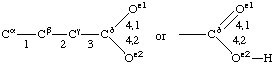 | Glutamine | 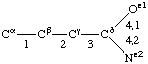 | ||
| Arginine | 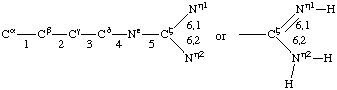 | ||||
| (c) Cyclic side chains | ||||
|---|---|---|---|---|
| Proline | 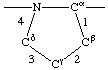 | | Hydroxyproline | 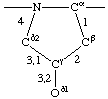 |
| Histidine | 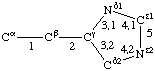 | Phenylalanine | 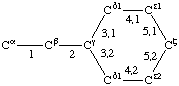 | |
| Tyrosine | 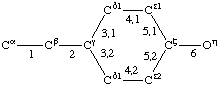 | Tryptophan | 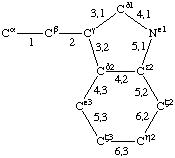 | |
If a side chain is branched, the branches are numbered 1 and 2, the order being determined (i) in cases where the branches are different, by application of Rule 2.2.1 or 2.3.1, (ii) in cases where two branches are identical (e.g., in valine, phenylalanine), by the application of Rule 2.2.2 (valine) or 2.3.2 (phenylalanine). Nonhydrogen atoms in different branches are designated by the Greek letter indicating their degree of remoteness from Cα and by the number of their branch (see Rules 2.2 and 2.3); e.g., in valine Ciγl and Ciγ2 (or Cγl(i ), Cγ2(i )). The branch number need not be indicated where no ambiguity results, e.g., in threonine Oγ and Cγ instead of Oγ1 and Cγ2, in hydroxyproline Oδ, Cδ instead of Oδ1, Cδ2, and in histidine Cδ, Nε, etc., instead of Cδ2, Nε2. For asparagine or glutamine, in cases where nitrogen and oxygen in the amide group have not yet been distinguished, these atoms may be written (NO)δ1, (NO)δ2, or (NO)ε1, (NO)ε2, the indices 1 and 2 being determined by Rule 2.3.2.
4.4. Designation of hydrogen atoms.
Hydrogen atoms are designated by the Greek letter and/or number of the atom to which they are attached, e.g., in valine Hiβ (or Hβ(i )). Where three hydrogen atoms are attached to a single non-hydrogen atom, they are designated 1, 2, and 3; in the situation the hydrogen atom giving the smallest (positive or negative) value of the principal torsion angle is designated 1, and the others are numbered in a clockwise sense when viewed in the direction B![[arrow right]](../greek/ARROWR.GIF) C (see Rule 2.2.3, which also covers the case where θ = ±60°), e.g., in valine Hiγ11, Hiγ12, Hiγ13 and Hiγ21, Hiγ22, Hiγ23, (or Hγ11(i ), etc.). Where only two hydrogen atoms are present, they are designated in accordance with Rule 2.2.2, case I for -CH2-R and case III for -NH2.
C (see Rule 2.2.3, which also covers the case where θ = ±60°), e.g., in valine Hiγ11, Hiγ12, Hiγ13 and Hiγ21, Hiγ22, Hiγ23, (or Hγ11(i ), etc.). Where only two hydrogen atoms are present, they are designated in accordance with Rule 2.2.2, case I for -CH2-R and case III for -NH2.
4.5. Designation of bonds and torsion angles (See Table III).
4.5.1. Bonds are designated by means of tho two atoms terminating them, e.g., Ciα-Ciβ, Niζ-Hiζ2, or, if no ambiguity results, by the symbol of the first atom of the bond, e.g., Ciα, Ciγ1. In superscripts the bond may be denoted either by α; β; γ1; γ2, etc., or by 1; 2; 3,1; 3,2; etc. Bond lengths are denoted b(Ciα, Ciβ), bCiα, b11, bi3.1, etc.
4.5.2. Torsion angles are denoted by X and are specified by two (or three) superscripts, the first one (or two), in the situation 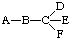 indicating the bond B-C about which the angle is measured, and the last indicating whether the angle is measured relative to D, E, or F. The principal torsion angle is defined by Rule 2.2.1, and if there is no ambiguity the last superscript may be omitted in referring to it.
indicating the bond B-C about which the angle is measured, and the last indicating whether the angle is measured relative to D, E, or F. The principal torsion angle is defined by Rule 2.2.1, and if there is no ambiguity the last superscript may be omitted in referring to it.
Thus in valine χi2,1 and χi2,2 refer to the torsion angles specifying atoms Ciγ1 and Ciγ2; in leucine χi3,1,1 χi3,1,2 and &chii3,1,3 refer to the torsion angles specifying the three hydrogen atoms attached to Cδl. If there is no ambiguity the principal torsion angles may be referred to, in valine and leucine, as χi2 and χi3,1, respectively. Corresponding notations without subscripts are χ2,1(i ), χ2(i ); χ3,1,1(i ), χ3,1(i ).
Note. By the sequence rule, when χ1 = 0, Cγ (or Cγ1) is in the eclipsed position relative to N.
5.l. Polarity of hydrogen bonds.
In specifying a hydrogen bond as directed from residue i to residue k (or from atom Xi to atom Yk), the direction X-H to :Y is implied; i.e, the atom covalently linked to the hydrogen atom is mentioned first.
Example
In the α helix the N-H of residue i is hydrogen bonded to the O=C of residue (i-4). Therefore, the α helix is described as having i to (i-4), or (5-1), hydrogen bonding.
5.2. Dimensions of hydrogen bonds.
Dimensions may be denoted by natural extensions of the nomenclature given above. For example, in 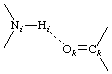 the following symbols might be used: b(Hi, Ok), τ(Ni,Hi,Ok), τ(Hi,Ok,Ck), θ(Hi,Ok), θi(N,H), θk(C,O).
the following symbols might be used: b(Hi, Ok), τ(Ni,Hi,Ok), τ(Hi,Ok,Ck), θ(Hi,Ok), θi(N,H), θk(C,O).
A regular helix is strictly of infinite length, with all φ's identical and all ψ's identical. A helical segment of polypeptide chain may be defined either in terms of φ and ψ or in terms of symmetry and hydrogen-bond arrangement.
In the description of helices or helical segments the following symbols should be used: n=number of residues per turn; h=unit height (translation per residue along the helix axis); t=360°/n=unit twist (angle of rotation per residue about the helix axis).
6.2. Definition in terms of φ and ψ.
Under this definition a helical segment is referred to as a (φ,ψ) helix; thus a right-handed α helix would be a (-57°,-47°) helix. The first and last residues of the helical segment are taken to be the first and last residues which have φ and ψ values equal to those defining the helix, within limits which should be defined in the context. No account is taken of hydrogen-bonding arrangements.
6.3. Definition in terms of symmetry and hydrogen-bond arrangement.
A helix is referred to as an nr helix, where n = number of residues per turn and r = number of atoms in ring formed by a hydrogen bond and the segment of main chain connecting its extremities. Thus an α helix would be 3.613. The first helical residue is taken as the first whose CO group is regularly bonded to NH along the helix (in the case of an α helix, to the NH of the fifth residue); the last helical residue is the last whose NH is regularly hydrogen bonded to CO along the helix (in the case of an α helix, to the CO of the residue last but four). Irregular hydrogen-bonding arrangements are not considered to form part of the helix.
Notes.
(i) A helical segment defined by Rule 6.2 may, but need not necessarily, be two residues shorter than the same segment defined by Rule 6.3.
(ii) These rules prescribe no definitions for irregular helical segments.
Recommendation A. Conformation and Configuration
There is at present no agreed definition of these two terms for general stereochemical usage.
In polypeptide chemistry the term "conformation" should be used, in conformity with current usage, to describe different spatial arrangements of atoms produced by rotation about covalent bonds; a change in conformation does not involve the breaking of chemical bonds (except hydrogen bonds) or changes in chirality (see Cahn et al., 1966).
On the other hand in polypeptide chemistry the term "configuration" is currently used to describe spatial arrangements of atoms whose interconversion requires the formal breaking and making of covalent bonds (note: this usage takes no account of the breaking or making of hydrogen bonds). For a more extensive discussion see IUPAC Tentative Rules for the Nomenclature of Organic Chemistry, Section E, Fundamental Stereochemistry, IUPAC Information Bulletin No. 35, 71-80 (1969).
Recommendation B. Definitions of Primary, Secondary, Tertiary, and Quaternary Structure
These concepts, originally introduced by Linderstrøm-Lang (1952), cannot be defined with precision, but the definitions given below may be helpful.
The use of the terms "primary, secondary, tertiary , and quaternary structure" has been criticized as being imprecise by Weltaufer (1961). He has proposed all alternative terminology.
The primary structure of a segment of polypeptide chain or of a protein is the amino acid sequence of the polypeptide chain(s), without regard to spatial arrangement (apart from configuration at the α-carbon atom).
Note. This definition does not inc1ude the positions of disulfide bonds and is therefore not identical with "covalent structure."
The secondary structure of a segment of polypeptide chain is the local spatial arrangement of its main-chain atoms without regard to the conformation of its side chains or to its relationship with other segments.
The tertiary structure of a protein molecule, or of a subunit of a protein molecule, is the arrangement of all its atoms in space, without regard to its relationship with neighboring molecules or subunits.
The quaternary structure of a protein molecule is the arrangement of its subunits in space and the ensemble of its intersubunit contacts and interactions, without regard to the internal geometry of the subunits.
Note. A protein molecule not made up of at least potentially separable subunits (not connected by covalent bonds) possesses no quaternary structure. Examples of proteins without quaternary structure are ribonuclease (one chain) and chymotrypsin (three chains).
Arnott, S., and Dover, S. D. (1967), J. Mol. Biol. 30, 209.
Arnott, S., and Dover, S. D. (1968), Acta Cryst. B24, 599.
Arnott, S., Dover, S. D., and Elliott, A. (1967), J. Mol. Biol. 30, 201.
Cahn,R.S.(1964), J. Chem. Educ. 41, 116.
Cahn, R. S., and Ingold, C. K. (1951), J. Chem. Soc. 612.
Cahn, R. S., Ingold, C. K., and Prelog, V. (1956), Experientia 12, 81.
Cahn, R. S., Ingold, C. K., and Prelog, V. (1966), Angew. Chem., Int. Ed. Engl. 5, 385, 511; Angew. Chem. 78, 413.
Edsall, J. T., Flory, P. J., Kendrew, J. C., Liquori, A. M., Némethy, G., Ramachandran, G. N., and Scheraga, H. A. (1966), J. Biol. Chem. 241, 1004; Biopolymers 4, 130; J. Mol. Biol. 15, 339.
Eliel, E. L. (1962), Stereochemistry of Carbon Compounds, McGraw-Hil1, New York, N. Y., pp 92-94.
IUPAC Information Bulletin (1969), No. 35, 36-80.
Klyne, W., and Prelog, V. (1960), Experientia 16, 521.
Linderstrøm-Lang, K. (1952), Proteins and Enzymes, Lane Memorial Lectures, Stanford University Press, Stanford, Calif., p 54.
Ramachandran, G. N., Ramakrishnan, C., and Sasisekharan, V. (1963), J. Mol. Biol. 7, 95.
Ramachandran, G. N., and Sasisekharan, V. (1968), Advan. Protein Chem. 23, 283.
Ramachandran, G. N., Sasisekharan, V., and Ramakrishnan, C. (1966), Biochim. Biophys. Acta 112, 168.
Schellman, J. A., and Schellman, C. (1964), Proteins 2, 1.
Traub, W., and Schmueli, U. (1963), in Aspects of Protein Structure, Ramachandran, G. N., Ed., Academic Press, London, p 81.
Wetlaufer, D. B. (1961), Nature 190, 1113.
Yonath, A.,and Traub, W. (1969), J. Mol. Biol. 43, 461.